エルサ・パンチローリ 著 的場和之 訳「哺乳類前史」メモ

エルサ・パンチローリ 著 的場和之 訳
「哺乳類前史」メモ第3章 頭にあいた穴ひとつ
----------------------------------------------------------------------------------
【まとめ】
・3億6千万年~3億年前の石炭紀、無脊椎動物を追い、水からでた最初の四肢動物は、背骨と4本の脚をもつすべての動物たち(哺乳類、両生類、爬虫類、鳥類)の共通祖先。
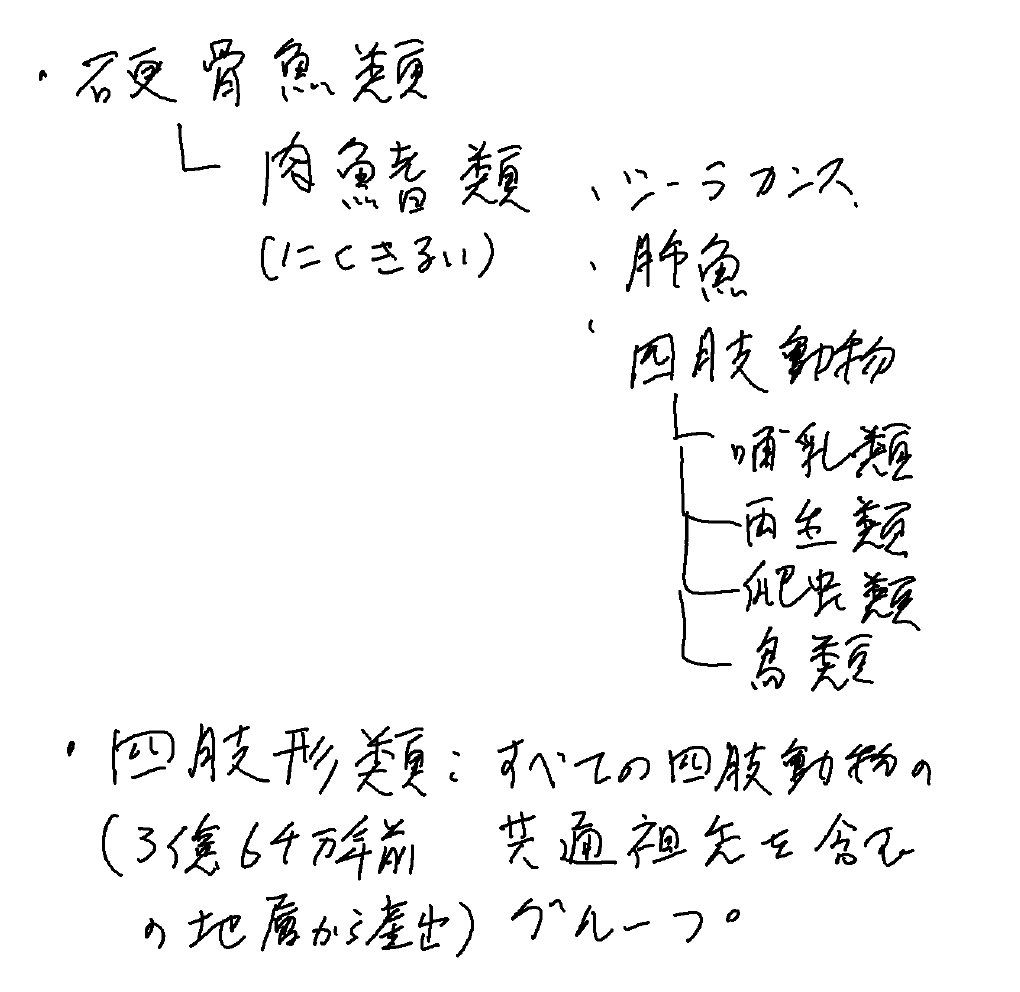
・四肢動物の祖先たちは完全な水中生活者で、硬骨魚類のなかの肉鰭類(にくきるい)と呼ばれるグループで(シーラカンス、肺魚、四肢動物)、ヒトはきわめて派生的な魚。
・陸上脊椎動物の二大系統の祖先は有羊膜類と無羊膜類で、有羊膜類の分類の二大系統は、単弓類と竜弓類(爬虫類)で、哺乳類は単弓類に属す。
----------------------------------------------------------------------------------
・現代の大気の酸素濃度は20%ほど。
・3億6千万年~3億年前の石炭紀は、大気の1/3が酸素だった。
・最初の森林は、シダ、コケ、トクサの巨大な祖先や親戚で構成された。
・現代の「樹」は進化的には新参者。
・石炭紀前半のパンゲアの森林は、小葉植物の天下だった。
・高さ30mを越え、産業革命を支えた石炭の大部分は、こうした植物の落ちた枝。
・4億7千万年前のオルドビス紀に菌類、植物が大地に進出。
・5000万年後、節足動物が続いた。
・節足動物は7000万年繁栄し、その後背骨のある動物が陸へ進出。
・節足動物:体節、硬い外骨格、関節のある付属肢をもつ、すべての動物の総称
・昆虫、甲殻類、クモ形類、多足類などを含む。
・かれらは急速にたくさんの形態に進化。
・クモとヤスデは先駆者のなかの先駆者。
・空気呼吸の最初の化石証拠は、4億2300万年前の地層から見つかったヤスデであるニューモデスムス。
・外骨格に気門と呼ばれる小孔が空いていた。
・気門は体内の血管や臓器とつながり、ガス交換(酸素を取り入れ二酸化炭素を排出)を担う。
・気門は収斂進化により節足動物の複数のグループで独立に獲得され、今日まで維持されている。
・気門がたくさんあると、水分も漏れ出してしまう。
→昆虫や多足類が小さい理由
・大きく成長するにはたくさんの酸素が必要。
・気門を大きく多くしなければならないが、やりすぎると干からびて死ぬ。
・現在の昆虫の最大体長の上限は約15センチメートル。
・節足動物の体のサイズの上限は、大気に占める酸素の割合が高ければ変わる。
・当時の昆虫は現在よりもはるかに巨大だった。
・現生種の10倍サイズの巨大昆虫も石炭紀には珍しくなかった。
・石炭紀の湿地林で、最初の四肢動物は無脊椎動物を追い、水からでた。
・彼らは、背骨と4本の脚をもつすべての動物たち(哺乳類、両生類、爬虫類、鳥類)の共通祖先。
・四肢動物の祖先といえる生き物たちは、完全な水中生活を送っていた。
・硬骨魚類のなかの肉鰭類(にくきるい)と呼ばれるグループ。
・シーラカンス、肺魚、四肢動物は肉鰭類を構成。
・ヒトはきわめて派生的な魚。
・四肢形類:すべての四肢動物の共通祖先を含むグループ
・最初期のメンバーは、デボン紀の地層(約3億6千万年前)から産出。
・われわれの先祖は、アカントステガに似た動物だったはず。
・ヒトの腕の長さくらいのサンショウウオに似た生物。
・扁平な頭をもち、眼は頭頂部にあり、突き出した四肢の先から、パドルのように幅広の指が伸び、尾は方向舵のように扁平。
・四肢や指は、肉鰭類たちが水から出られるように発達したわけではない。
・進化は最終目的を念頭に進むわけではない。
・デボン紀末、3億7500万年前~3億6千万年前にかけて二度の大量絶滅が地球上の生命を激減させた。
・その後1500万年の間、化石記録は沈黙を保った。
・この後、四肢動物は水の外でも自重を支えられる陸上生活者として多様化をとげた。
・外適応:ある目的のために進化した形質が、ほかの目的のために転用さされる。
・四肢動物の祖先が陸上生活のために適応を進化させたことはない。
・すでに備えていた適応が、たまたま好都合だったおかげで、それらを利用して陸上に進出できた。
・哺乳類の物語の始まりにふさわしい動物:ウエストロシアナ
・最初期の陸上生物のひとつだっただけでなく、ヒトが属するグループの有羊膜類との類縁関係が見てとれる。
・陸上脊椎動物の二大系統の祖先
有羊膜類
無羊膜類
・無羊膜類:水に頼って繁殖、水分を保ち酸素を与えてくれる水の中に卵を産む。→カエル、サンショウウオ、アシナシイモリ
・羊膜:発生途中の胚を包み込む、液体で満たされた膜の名前。カエルの卵塊のようなゼリー状の表層が進化したもの。
・両生類の場合、このゼリー状物質が卵と周囲の水の間でおこなわれる老廃物の排出や交換を担う。
・有羊膜類では、同じ機能を、羊膜が外で実現している。
・羊膜は発生途中の胚を包む膜のクッションを形成、そのなかに持ち運び可能な池である羊水を満たす。
・卵殻がこれらすべてを内包する。→両生類と魚類の卵にはない特徴
・卵が獲得したイノベーションのおかげで、有羊膜類は水辺を離れて子育て可能になった。
・水から出た脊椎動物は、水中より30倍も酸素を多く含む大気を利用できた。
・最初の四肢動物は「口腔ポンピング」を採用した。
・口腔の上げ下げで生じる、ふいごのようなメカニズム。
・初期の四肢動物の多くが幅広く平たい頭部をもっていた理由。
・ぺしゃんこな形は広い口腔をつくりだす。
・最初の四肢動物はみな口呼吸をしていた。
・有羊膜類は口腔ポンピング→胸の筋肉を使う胸式呼吸へ変化した。
→有羊膜類はより効率よく呼吸できるようになり、直立した姿勢と長い首というプロポーションが可能になった。
・頭骨と顎の筋肉の一部を新たな採食様式に転用できた。
→顎の先端部分でかじり取る技術が必要な、植物食が初めて実現
・共有派生形質:あるグループと別のグループを区別する基準
・有羊膜類の共有派生形質
①骨の配置:前頭骨の幅が広く、眼窩の一部を形成。
②口蓋:口腔の天井部分である口蓋が、喉の奥の方へ向かい、歯がずらりと並んだ縁に囲まれている。
③肩の骨がより複雑になっている。(陸上生活ともなう前肢の使い方の変化に関係するもの)
④くるぶしと手首は簡素化し、複数の骨が癒合して距骨(きょこつ、くるぶしの一部)を形成。
・約3億年前、哺乳類の系統はいとこである爬虫類と袂を分けていた。
・哺乳類が爬虫類から進化した、というのは「誤解」。
・哺乳類と爬虫類には共通の祖先がいた。
・最初の羊膜をもつ四肢動物は、哺乳類でも爬虫類でもなかった。
・どちらのグループもまだ進化していなかった。
・石炭紀、ヒトとカメ、ワニ、恐竜、鳥、トカゲの最後の共通祖先は役目を終え、消えていった。
・有羊膜類の分類の二大系統
単弓類
竜弓類
・竜弓類には、すべての哺乳類やさまざまな絶滅動物が含まれる。
・竜弓類(爬虫類)は、四肢動物一族のなかで最大の成功者。
カメ、翼竜、トカゲ、ムカシトカゲ、魚竜、ワニ、恐竜
・恐竜の現生の子孫である鳥は、哺乳類の2倍の種数を誇る。
・最初の単弓類、哺乳類とその親戚からなる四肢動物の系統の創始者の候補は、ノバスコシア州の石炭紀の地層から発見された。
・哺乳類の系統に属すると考えられる化石のうち、最近見つかった化石は、アサフェステラ、プロトクレプシドロプス、アルカエオティリス、エチネルペトン。
・アサフェステラの小さく幅の広い頭骨が、初期単弓類に特有のパターンで構成されている。
・これが確かなら、アサフェステラは最古の幹哺乳類のひとつといえる。
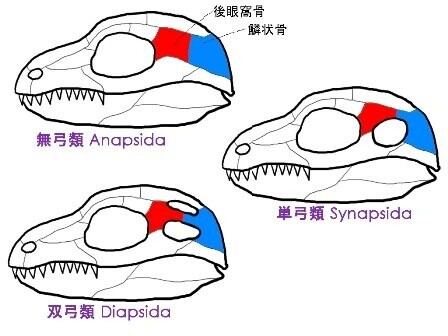
・単弓類がもつ決定的特徴:頭骨の両側にひとつずつ、側頭窓と呼ばれる穴をもつ
・側頭窓は眼のすぐうしろにあり、すべての単弓類において左右一つずつ。
・側頭窓には筋肉が通り、この穴のおかげで口を開閉する筋肉が付着する部分が確保されている。
・爬虫類の系統にみられる頭骨の穴の数のパターンは複雑。
・穴の獲得と喪失が複数のグループで起こったから。
・ほとんどの爬虫類は穴ふたつの双弓類だが、カメは穴のない無弓類。
・単弓類はほぼ例外なく単一の側頭窓をもつ。
・単弓類と竜弓類の最大の違いにあいまいな部分はなく、頭の両側にひとつずつあいた穴は、わたしたちを定義づける一貫した特徴。
・初期有羊膜類、単弓類と竜弓類を「爬虫類的」と呼ぶのは正しくない。
・現生爬虫類を「初期有羊膜的」と形容すべき。
・現生爬虫類の分類群、とくにトカゲは先祖を彷彿させる数多くの特徴を維持。
・表面的にはあまり変わっていないレトロなグループ。
・哺乳類は劇的に、明白に変化した。
・哺乳類が爬虫類から進化したのではない。
・初期単弓類の爬虫類っぽさは、爬虫類との共通祖先である、初期有羊膜類のボディプランの名残にすぎない。
・哺乳類と爬虫類、単弓類と竜弓類の間の断絶は深く、その端緒は石炭紀にさかのぼる。